Evolutietheorie en organisatiestrategie
Het belang van evolutionaire processen voor de strategieontwikkeling
nothing makes sense except in the light of evolution
(vrij naar Dobzhansky, geciteerd in Evolutie 1998, cursusdeel 1: 7)
3 Evolutie
Dit hoofdstuk beschrijft hoe uit de literatuur over evolutie het algemene model voor evolutie is afgeleid, welke evolutionaire processen zijn geïdentificeerd, en welke voorwaarden voor effectieve ontwikkeling op basis daarvan zijn geformuleerd. Daartoe wordt in § 3.1 de ontwikkeling in het denken geschetst die loopt van Darwin, via Donald T. Campbell naar Weick. Darwin omschreef natuurlijke selectie in een biologische context. Campbell verbreedde dit tot een algemeen, van de biologische context geabstraheerd, model. Weick gebruikt Campbell’s model bij de bestudering van processen waarin mensen hun handelen en denken organiseren, anders gezegd: in de context van sociaal-psychologische processen van organiseren. Paragraaf 3.1 wordt afgesloten met een eerste globale beschrijving van het algemene model van evolutie. Paragraaf 3.2 gaat dieper in op de begrippen ‘variatie’, ‘selectie’ en ‘retentie’ die tezamen de basis voor het algemene model vormen, en wordt afgesloten met de behandeling van de effecten van verwevenheid en aggregatie op het model. In § 3.3 wordt het algemene model voor evolutie uitgewerkt, waarna in § 3.4 de voorwaarden voor effectieve ontwikkeling zoals die naar voren gekomen zijn in de bespreking van de deelprocessen variatie, selectie en retentie, worden gerelateerd aan het algemene model.
3.1 Darwin, Campbell, Weick
In zijn in 1859 gepubliceerde boek The origin of species gaf Darwin een nog steeds adequate beschrijving van het verschijnsel natuurlijke selectie in een biologische context [1]:
As many more individuals of each species are born than can possibly survive; and as, consequently, there is a frequently recurring struggle for existence, it follows that any being, if it vary however slightly in any manner profitable to itself, under the complex and sometimes varying conditions of life, will have a better chance of surviving, and thus be naturally selected. From the strong principle of inheritance, any selected variety will tend to propagate its new and modified form. (1859: 68).
Ten opzichte van Darwin’s oorspronkelijke beschrijving is de enige grote verandering in de biologische evolutietheorie het toegenomen inzicht in de wijze waarop variatie tot stand komt en wordt doorgegeven aan volgende generaties. Voor Darwin was evolutie descent with modification, afstamming gepaard met modificatie, maar hij wist niet hoe die modificatie, die variatie, tot stand komt, noch hoe deze wordt doorgegeven. Pas tussen 1930 en 1940 werd de evolutietheorie uitgebreid met ideeën uit de genetica, en vanaf die periode wordt de theorie ‘neodarwinistisch’ genoemd (Evolutie 1998, cursusdeel 1: 16, 22, 50).
Tussen 1950 en 1960 vond er een opleving plaats in de belangstelling voor sociale evolutie (Campbell 1965: 19). De sociaal-psycholoog Campbell [2] is een van diegenen die een brug heeft geslagen tussen de biologische evolutieleer zoals die zich tot op dat moment had ontwikkeld, het neodarwinisme dus, en de sociale wetenschappen. [3]
Onder ‘evolutie’ wordt in dit rapport verstaan de van de biologische context geabstraheerde betekenis van natuurlijke selectie, genetische drift en mutatie. Evolutionaire processen worden beschouwd als doelloos, en ontwerp als een interpretatie achteraf van het resultaat van selectie. Dit laat geen ruimte voor determinisme – de toekomst ligt niet vast – en voor waardeoordelen over het resultaat van het proces – het Panglossianisme van Spencer was een historische vergissing. Binnen deze aan het begrip ‘evolutie’ toegekende betekenis is in het onderzoek de uitwerking daarvan gehanteerd zoals Campbell die formuleerde onder de noemer blind variation and selective retention (BVSR):
The three essentials are these:
1. The occurence of variations: heterogeneous, haphazard, “blind”, “chance”, “random”, but in any event variable. (The mutation process in organic evolution, and exploratory responses in learning).
2. Consistent selection criteria: selective elimination, selective propagation, selective retention, of certain types of variations (Differential survival of certain mutants in organic evolution, differential reinforcement of certain responses in learning).
3. A mechanism for the preservation, duplication, or propagation of the positively selected variants (the rigid duplication process of the chromosome-gene system in plants and animals, memory in learning).
Given these conditions, an evolution in the direction of better fit to the selective system becomes inevitable. (ib.: 27).
Enerzijds is Campbell geïnspireerd door het model voor natuurlijke selectie zoals geformuleerd door Darwin, anderzijds beschouwt hij biologische evolutie als een speciaal geval van een algemeen model: ‘The analogy (…) will not be from organic evolution per se, but rather from a general model for adaptive fit or quasiteleological processes for which organic evolution is but one instance.’ (ib.: 26). Het ‘general model’ waarnaar Campbell in het voorgaande citaat verwijst staat centraal in Campbell’s visie op Popper’s kennisleer. Campbell (1974c) betitelt deze als evolutionary epistemology (evolutionaire kennisleer). [4] Campbell’s dogma [5] stelt dat elke toename in fit tussen systeem en omgeving is gebaseerd is op BVSR-processen, en dat de vele processen die een volledig BVSR-proces omzeilen door gebruik te maken van reeds opgebouwde kennis van de omgeving, zelf het resultaat zijn van BVSR-processen:
A. A blind-variation-and-selective-retention process is fundamental to all inductive achievements, to all genuine increases in knowledge, to all increases of fit of system to environment.
B. In such a process the are three essentials: (1) Mechanisms for introducing variation; (2) Consistent selection processes; (3) Mechanisms for preserving and/or propagating the selected variations. Note that in general the preservation and generation mechanisms are inherently at odds, and each must be compromised.
C. The many processes which shortcut a more full blind-variation-and-selective-retention process are in themselves inductive achievements, containing wisdom about the environment achieved originally by blind variation and selective retention.
D. In addition, such shortcut processes contain in their own operation a blind-variation-and-selective-retention process at some level, substituting for overt locomotor exploration or the life-and-death winnowing of organic evolution. (ib.: 421).
Dat Campbell een inspiratiebron voor velen is geweest, getuigt het boek Variations in organization science: In honor of Donald T. Campbell (Baum & McKelvey 1999). Een van degenen die door Campbell werd geïnspireerd was de sociaal-psycholoog Weick. [6] Weick beschrijft in zijn boek The social psychology of organizing de analogie tussen sociaal-psychologische processen van organiseren en de processen die een rol spelen bij natuurlijke selectie: ‘Organizing processes resemble closely the three processes commonly associated with theories of natural selection.’ (Weick 1979: 119). Weick baseert zijn analogie op het BVSR-model van Campbell, en neemt in zijn model niet alleen de drie processen variatie, selectie en retentie op, maar ook de bron van variatie, ecological change. Weick gebruikt overigens het begrip enactment in plaats van het begrip ‘variatie’ zoals dat door Campbell wordt gebruikt. Het volgende citaat verduidelijkt de positie die Weick toekent aan ecological change:
Within de flows of experience that engage people and activities there frequently are changes or differences. (…) These differences may provide the occaision for attempts at equivocality removal and attempts to determine the significance or triviality of the differences. Ecological changes provide the enactable environment, the raw materials for sense-making. (…) This portion of the organizing process is dubbed ecological change to capture the fact that people normally are not aware of things that run smoothly. It is only the occasion of change when attention becomes active. (ib.: 130).
Het door Weick gebruikte begrip enactment brengt tot uitdrukking dat mensen door te handelen begrip krijgen van hun omgeving, ‘(…) enactment is first and foremost about action in the world (…)’ (Weick 1995: 36). Weick heeft het begrip enactment ontleend aan de juridische context: to enact betekent bepalen, een enactment is een verordening, een wet. ‘I like the word because it suggests that there are close parallels between what legislators do and what managers do. Both groups construct reality through authoritative acts. When people enact laws, they take undefined space, time, and action and draw lines, establish categories and coin labels that create new features of the environment that did not exist before.’ (ib.: 30-31). Het volgende citaat illustreert het belang van Weick’s visie voor de strategieontwikkeling: hij ziet organisatie als de uitkomst van evolutionaire processen van handelen en waarnemen, waarbij de veranderde omgeving een deel is van die uitkomst:
I use the word enactment to preserve the fact that, in organizational life, people often produce part of the environment they face (…) There is not some impersonal “they” who puts these environments in front of passive people. Instead, the “they” is people who are more active. All too often people in organizations forget this. They fall victim to this blindspot because of an innocent sounding phrase, “the environment”. The word the suggests something that is singular and fixed; the word “environment” that this singular, fixed something is set apart from the individual. Both implications are nonsense.’ (ib.: 30-32).
Figuur 3-1, ontleend aan Weick (1979: 132), geeft het globale model weer zoals Weick dat beschrijft. (Het is een causaal model: de +-tekens geven een versterkend effect in de richting van de pijl aan, terwijl het (+, –)-teken aangeeft dat er in de richting van de pijl, afhankelijk van de situatie, zowel versterking als demping kan plaatsvinden.)

Figuur 3-1 Weick’s enaction, selection, retention-model
De relaties tussen het biologische evolutiemodel, het BVSR-model van Campbell en het model van Weick zijn weergegeven in Figuur 3-2: het model van Campbell is geïnspireerd door het model van biologische evolutie en het op het model van Campbell gebaseerde model van Weick is op zijn beurt geplaatst in de specifieke context van sociaal-psychologische organiseerprocessen.
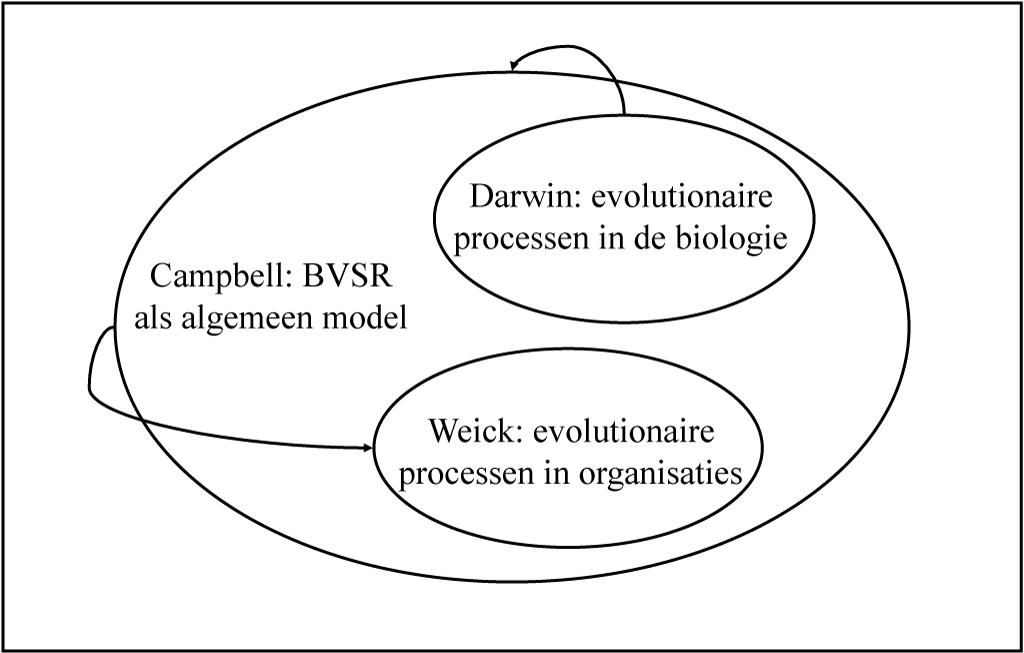
Figuur 3-2 De relatie tussen de theorieën van Darwin, Campbell en Weick
Paragraaf 3.2 gaat dieper in op variatie, selectie en retentie. Dit gebeurt tegen de achtergrond van de inzichten van Campbell en Weick. Figuur 3-3 geeft het op deze inzichten gebaseerde model weer zoals dat in dit onderzoek is gebruikt. Het evolutionaire proces is opgebouwd uit drie deelprocessen: variatie, selectie en retentie. Systeem en omgeving staan via het evolutionaire proces met elkaar is wisselwerking. In het systeem zelf en in de omgeving treden gebeurtenissen op die, via het variatieproces, leiden tot variaties. Deze variaties worden op basis van selectiecriteria uit het systeem zelf en uit de omgeving al dan niet geëlimineerd door het selectieproces. De variaties die niet geëlimineerd worden, worden door het retentieproces vertaald in veranderingen die deel uit gaan maken van het systeem en van de omgeving.

Figuur 3-3 De schematische weergave van het algemene model van evolutie
3.2 Natuurlijke selectie: variatie, selectie, retentie
In deze paragraaf worden achtereenvolgens variatie, selectie en retentie beschreven. De opdeling van het evolutionaire proces in drie subprocessen, waarmee gesuggereerd wordt dat deze volgtijdelijk plaatsvinden, is vanuit analytisch oogpunt te verdedigen: zonder deze opdeling is een gedetailleerde analyse immers niet goed mogelijk. Daarbij dient echter in gedachten gehouden te worden dat deze processen in werkelijkheid vaak sterk verweven zijn. Dit komt tot uitdrukking in begrippen zoals selective retention in de titel van Variation and selective retention in socio-cultural evolution (Campbell 1965). Het laatste deel van deze paragraaf gaat dieper in op deze verwevenheid en op de begrippen ‘aggregatie’ en ‘hiërarchische gelaagdheid’.
Variatie
variations, a heterogeneity of alterations on existing form
(Campbell 1974b: 143)
Campbell omschreef zijn model oorspronkelijk als blind-variation-and-selective-retention, waarbij het woord blind uitdrukt dat het resultaat van een variatie niet vooraf bekend mag zijn (1965: 28). Blinde, willekeurige variatie is het alternatief voor variatie waarvan vooraf wordt vastgesteld dat deze zinvol is. Dit ‘zinvol’ impliceert dat op voorhand kan worden vastgesteld dat deze variatie tot wenselijke resultaten leidt; deze gedachtegang gaat voorbij aan de conclusie van de analyse van Campbell dat de aanwezigheid van blinde variatie een voorwaarde is voor werkelijk nieuwe inzichten. Later stelde Campbell in plaats van blind het begrip unjustified voor (1974b: 151). Met dit begrip bracht het risico van het veronderstellen van voorkennis van het resultaat van een variatie expliciet onder woorden. Hoewel unjustified beter dan blind onder woorden brengt welke eis Campbell aan variatie stelt, is het model bekend onder de naam blind-variation-and-selective-retention.
Nelson en Winter stellen zich in hun boek An evolutionary theory of economic change (1982) op het standpunt dat het onderscheid tussen ‘blind’ en ‘deliberate’ zinloos is in de context van hun theorie: ‘(…) some people who are particularly alert to teleological fallacies in the interpretation of biological evolution seem to insist on a sharp distinction between explanations that feature the processes of “blind” evolution and those that feature “deliberate” goal-seeking. Whatever the merit of this distinction in the context of the theory of biological evolution, it is unhelpful and distracting in the context of our theory of the business firm.’ (ib.: 10-11). Bij dit citaat kunnen twee kanttekening worden geplaatst. Ten eerste dat Nelson en Winter blind evolution-processen plaatsen tegenover deliberate goal-seeking-processen, maar verder niet op deze begrippen ingaan. Zij ontkennen (onderkennen?) daarmee niet het pleonastische karakter van de uitdrukkingen blind evolution en deliberate goal-seeking maar volstaan met te benadrukken dat het onderscheid voor hen niet relevant is. Daarmee geven zij aan dat ze het begrip ‘evolutie’ gebruiken in de betekenis geleidelijke ontwikkeling en nemen ze afstand van het begrip ‘natuurlijke selectie’. Ten tweede gaan Nelson en Winter een eventuele discussie uit de weg door hun visie, ‘it is unhelpful and distracting in the context of our theory’, niet te onderbouwen. Dat maakt het onmogelijk hun oordeel over de door Campbell geformuleerde eis van unjustified variation te toetsen. De verwijzing naar Campbell door Nelson en Winter is echter positief: ‘Campbell (1969) [mijn verwijzing 1965] provided an excellent survey of that broad field (…)’ (ib.: 43, cursivering toegevoegd). Het dogma van Campbell (zie p. 13) kan wellicht een brug slaan tussen het afwijzen van het belang van blind variation en het positieve oordeel over het werk van Campbell als het gebruik van goal-seeking wordt gezien als het resultaat van een BVSR-proces.
Campbell verwoordt de essentie van de aan variaties gestelde blind-eis als volgt: ‘In going beyond what is already known, one cannot go but blindly. If one can go wisely, this indicates already achieved wisdom of some general sort.’ (Campbell 1974c: 422).
Een andere voorwaarde wordt door Campbell meer impliciet gesteld. Er moet sprake zijn van verschillende variaties omdat anders selectie niet mogelijk is. Uit één variatie kan niet worden geselecteerd, er is geen criterium waarmee een variatie kan worden vergeleken: ‘beter’ is een relatief begrip, het zegt iets over een variatie in vergelijking tot de andere variaties waaruit gekozen is. Deze redenering leidt tot een additionele voorwaarde die in navolging van Campbell door Romanelli aan de variaties wordt gesteld, namelijk dat deze ‘fouten’ bevat, te weten die variaties die in het selectieproces worden geëlimineerd: ‘Novel solutions can be revealed only after they have been tried. According to Campbell, human actors need to understand variations as the trials that they are, and evaluate them against outcomes instead of, more commonly, assuming them as solutions to be defended. In others words, failure must be possible.’ (1999: 84).
Dat een variatie fout is, wordt pas na het ontstaan van de variatie vastgesteld, namelijk in het selectieproces, wanneer de betreffende variatie níet wordt geselecteerd. Zonder fouten valt er niets te selecteren en vindt er dus, per definitie, geen evolutie plaats. Zonder verder in te gaan op de noodzaak van fouten, wordt hier volstaan met de constatering dat de kwalificatie ‘fout’ samenhangt met de rol van de human actors zoals genoemd in het voorgaande citaat van Romanelli, en dat deze kwalificatie, net als design, is toe te schrijven aan the hindsight of a selective system (zie het citaat in bijlage 2).
Weick (1979: 129) formuleert een additionele eis aan de variaties: ‘Notice that the additional requirement of persistence until natural selection can operate may shrink drastically the pool of mutations available in any system.’ (Weick gebruikt hier het begrip mutations in een betekenis die overeenkomt met wat hier variaties genoemd wordt, de uitvoer van het variatieproces). De variaties dienen dus lang genoeg te bestaan om te kunnen worden verwerkt door het selectieproces.
Voor het overige baseert Weick zich expliciet (ib.: 122) op het model van Campbell en refereert bij de beschrijving van variatie (ib.: 123) aan publicaties van Campbell (1965; 1974b; 1974c). Bij de beschrijving van zijn eigen model (zie Figuur 3-1 en het citaat dat daaraan voorafgaat) wordt het verschil tussen het model van Weick en dat van Campbell duidelijk. Weick neemt ecological change, en dus de omgeving, expliciet als element in zijn model op. Ook Campbell refereert frequent aan de omgeving, maar nergens op een wijze waarbij omgeving en variatie zo sterk samenhangen als bij Weick. Een deel van de verklaring hiervoor is Weick’s focus op psychologische en sociale processen: in zijn beschrijving van de relatie tussen ecological change en enactment klinkt het credo van de sociale wetenschappen door: ‘If men define situations as real, they are real in their consequences.’ (Thomas & Thomas 1928: 572); voor dit onderzoek is deze invalshoek relevant, want zeker voor het proces van strategieontwikkeling is er een ‘(…) active role (…) organizational members play in creating the environments which then impose on them.’ (Weick 1979: 130). Voor een ander deel kan het verschil worden verklaard door de verschillende manier waarop Campbell en Weick omgaan met de hiërarchische gelaagdheid van het systeem waarop hun modellen betrekking hebben. Deze gelaagdheid komt aan de orde aan het einde van deze paragraaf waar verwevenheid en aggregatie worden besproken.
Selectie
for certainly we do not yet know much about the
selective system that shaped civilised social man
(Campbell 1974a: 184)
Campbell gebruikt diverse begrippen die betrekking hebben op het selectieproces. Hij onderscheidt verschillende selective systems (1965: 29-31) en selective criteria (ib.: 31-33). In een latere publicatie gebruikt hij in plaats van selective criteria het begrip selectors (1974b: 145-147). Weick volgt Campbell in het gebruik van de begrippen selective systems (waaraan hij ook refereert als mechanisms) en selective criteria maar gebruikt, hoewel hij op diverse plaatsen refereert aan de genoemde publicatie van Campbell (1974b), nergens het begrip selector. In deze tekst wordt het begrip ‘selectiecriterium’ gebruikt in de betekenis die Campbell geeft aan selective criterion; Campbell’s selective systems blijven verder buiten beschouwing.
Campbell onderscheidt verschillende selectiecriteria: interne, die hij onderverdeelt in structurele en plaatsvervangende, en externe (Campbell 1974b: 145-147). Structurele selectiecriteria zijn wetmatigheden die gelden voor datgene, wat aan het selectieproces is onderworpen. Campbell geeft als voorbeelden de natuurwetten die het proces van kristalvorming in een verzadigde zoutoplossing en het proces van de vorming van sneeuwvlokken (mede) bepalen (ib.: 144-145). Dawkins noemt dit de selectie van stabiele vormen: ‘Darwin’s ‘survival of the fittest’ is really a special case of a more general law of survival of the stable. (…) The earliest form of natural selection was simply a selection of stable forms and a rejection of unstable ones.’ (1989: 12-13). Dennett gebruikt het begrip ‘gedwongen zetten’: ‘In dit soort gevallen is er om de een of andere reden maar één manier waarop de dingen gedaan kunnen worden. Maar redenen kunnen diep en oppervlakkig zijn. De diepere redenen zijn de beperkingen die worden opgelegd door de wetten van de natuurkunde (…)’ (1995: 126).
Plaatsvervangende selectiecriteria zijn interne representaties van externe selectiecriteria. Campbell geeft als voorbeeld de regeneratie van de poot van een salamander. De groei van de nieuwe poot lijkt door te gaan totdat deze de grond raakt. Wat er feitelijk gebeurt is dat interne selectiecriteria het groeiproces sturen en daarmee de lengte van de poot bepalen (Campbell 1974b: 146). Een voorbeeld van een plaatsvervangend selectiecriterium in bedrijfskundige context is die van durfkapitaalverschaffers: ‘Our understanding of entrepreneurial variation would be incomplete if we focused only on the direct selection environment for entrepreneurs. Venture capital is one of several vicarious selection systems that influence the degree of variety among new enterprises. Such systems help capitalist societies adapt rapidly to new technologies and markets by lowering the cost of experimentation.’ (Anderson 1999: 150). Weick geeft, overigens zonder dat als zodanig te benoemen, een voorbeeld van een plaatsvervangend selectiecriterium in de context van organisatiestructurering: ‘(…) decision makers in organizations intervene between the environment and its effects inside the organization, which means that selection criteria become lodged more in the decision makers than in the environment.’ (Weick 1979: 125).
Externe selectiecriteria zijn selectiecriteria uit de omgeving. ‘It is to this class of selectors that ‘fit’, ‘adaptation’, ‘correspondence’, ‘objectivity’ or ‘descriptive truth’ is due.’ (Campbell 1974b: 146). Weick besteedt weinig aandacht aan de selectiecriteria als zodanig en gaat niet expliciet in op het onderscheid intern, structureel, plaatsvervangend en extern. Hij besteedt wel aandacht aan het verschil tussen kunstmatige en natuurlijke selectie. Hij stelt dat selectieprocessen in een organisatie minder afhankelijk zijn van het toeval dan van de bewuste intenties van mensen: mensen in organisaties handelen meer als fokkers dan als gokkers: ‘The enacted environment is artificial rather than natural in the sense that it is laced with preferences, purposes, idiosyncratic punctuations, desires, selective perceptions, and designs. It is the environment of the breeder.’ (Weick 1979: 176). [7]
De effectiviteit van het selectieproces kan positief en negatief worden geformuleerd. Het selectieproces ontvangt als invoer variaties en moet de beste daarvan doorgeven aan het retentieproces. Negatief geformuleerd: een effectief selectieproces kenmerkt zich door de eliminatie - het niet-doorgeven aan het retentieproces - van mislukkingen. Positief geformuleerd: een effectief selectieproces kenmerkt zich door de propagatie - het doorgeven aan het retentieproces - van succesvol gebleken variaties. Campbell combineert de positieve en negatieve vorm: ‘Consistent selection criteria: selective elimination, selective propagation, selective retention (…)’ (1965: 27); ‘Systematic selection from among the variations. Systematic elimination.’ (1974b: 143).
De enige eis die door Campbell aan het selectieproces wordt gesteld is consistentie (zie de voorgaande citaten). Dit lijkt primair betrekking te hebben op consistentie in de tijd, maar Campbell beschrijft ook bedreigingen voor het selectieproces, met name voor de effectiviteit van de externe selectiecriteria, die de fit tussen systeem en omgeving in stand moet houden of verbeteren. Deze bedreigingen hangen nauw samen met de hiërarchische gelaagdheid van systemen en de rol die plaatsvervangende selectiecriteria daarin vervullen (zie het citaat van Weick op p. 18 over decision makers in organizations en de bespreking van verwevenheid en aggregatie aan het einde van deze paragraaf).
Retentie
retention means liability to recall, and it means
nothing more than such liability
(James, geciteerd in Weick 1979: 207)
Retentie in biologische systemen is concreet aanwijsbaar, retentie in sociaal-culturele systemen is daarentegen in veel gevallen moeilijk aanwijsbaar. Dat laatste zou zelfs reden kunnen zijn het idee van sociaal-culturele evolutie af te wijzen:
One of the things that impresses one in comparing biological evolution with socio-cultural evolution is the greater rigidity and loyalty of the biological retention system. Corresponding to this, no doubt, is the greater mutability and diversity of societal forms, so that in any modern society, by the time a total replacement of individuals has taken place (i.e., over the course of 50 to 100 years) the system has become a different “species,” so to speak.
This may, of course, merely be a function of our point-of-observation from within the current system. (…) Nonetheless, the absence of an effective retention system would be grounds for rejecting the notion of socio-cultural evolution. (Campbell 1965: 33-34).
De omschrijvingen die zowel Campbell als Weick geven van retentie zijn kort en liggen sterk in het verlengde van het selectieproces: dat wat geselecteerd is, wordt vastgelegd. Campbell (1965: 27) omschrijft dit als ‘A mechanism for the preservation, duplication, or propagation of the positively selected variants’, en geeft als voorbeeld daarbij de rol van het geheugen in het leerproces. Waar zowel Campbell als Weick veel aandacht aan besteden, is de wijze waarop retentie, met name in sociaal-psychologische en sociaal-culturele systemen plaatsvindt. Campbell noemt in een antropologische context gewoonten en rituelen (1965: 29; 1994: 28). Weick noemt in een organisationele context procedures en notulen en ook het geheugen van mensen, individueel of als groep.
Weick benoemt een aantal karakteristieken van retentie. Hij refereert daarbij aan een memory surface, een fysiek substraat waarin de herinnering wordt vastgelegd. De eerste karakteristiek betreft de kwaliteit van de registratie van de herinnering en benadrukt de verwevenheid die er kan bestaan tussen selectie en retentie. Weick illustreert dit met een citaat van De Bono:
The two ways in which a bad memory surface can be bad are distortion and incompleteness. (…) Incompleteness (…) is a tremendously important deficiency, for when some things are left out there must be some things which are left in. This implies a selecting process. And a selecting process is the most powerful of all information handling tools. It is quite likely that the great efficiency of the brain is not due to its being a brilliant computer. The efficiency of the brain is probably due to its being a bad memory surface. One could almost say that the function of the mind is mistake (De Bono, geciteerd in Weick 1979: 208).
De tweede karakteristiek die Weick benoemt, is de geheugenwerking van het memory surface. In zijn jelly model of retention komen nieuwe gegevens in de vorm van druppels heet water op een bodem van gelatine terecht. Dit hete water lost de gelatine voor een deel op, waardoor er op de plaats waar het water terechtkwam een kuiltje ontstaat. Het effect van de volgende druppels heet water is afhankelijk van het effect van de eerdere druppels, van de patronen van kuiltjes en verbindingen daartussen die als gevolg van eerdere ‘indrukken’ zijn ontstaan:
the memory surface does not accumulate memories bij simple addition. Since each memory is processed by the preceeding memories, the actual sequence of presentation may make a very great difference to the pattern that is established, even though the individual patterns presented may be the same (De Bono, geciteerd in Weick 1979: 211).
De constatering van De Bono en Weick dat natuurlijke selectie moet voortbouwen op dat, wat er als is, werd ook al gedaan door Darwin: ‘(…) throughout nature, almost every part of every being has probably served, in a slightly modified condition, for diverse purposes, and has acted in the living machinery of many ancient and distinct specific forms.’ (Darwin, geciteerd in Doolittle 1994: 59).
Verwevenheid en aggregatie
Retentie kan worden beschouwd als het laatste van een serie van drie deelprocessen. Deze drie deelprocessen kunnen echter ook worden beschouwd als delen van een continuproces waarbij die variaties die zijn geselecteerd door het selectieproces en vastgelegd door het retentieproces weer de basis vormen voor nieuwe variaties, en zo verder, ad infinitum. Zo weergegeven komt het spanningsveld tussen variatie en retentie nadrukkelijker in beeld, en wordt duidelijk dat deze processen elkaar tegenwerken. Campbell: ‘There is, perhaps, always a potential conflict between the freedom to vary, which makes advance possible, and the value of retaining the cultural cumulation.’ (1965: 35).
Het spanningsveld tussen variatie en retentie komt ook anders omschreven in de literatuur voor. Zo beschrijft March het als het spanningsveld tussen exploratie en exploitatie: ‘In evolutionary models of organizational forms and technologies, discussions of the choice between exploration and exploitation are framed in terms of balancing the twin processes of variation and selection.’ (1991: 72). Kauffman spreekt van de ‘struggle to invent and improve.’ (1995: 119). Ook Baaij en Van den Bosch signaleren dit spanningsveld, en zij leggen een verband tussen extern succes en interne inertie: ‘Efficiëntie en effectiviteit (exploitation) kunnen daarbij [bij extern succes] toenemen. De flexibiliteit en innovatie (exploration) zullen echter afnemen en de onderneming wordt relatief inert.’ (1999: 56; zie ook Volberda 1998; zie voor een bredere analyse van het effect van succes ook Miller 1999). Extrapolatie van deze invalshoek leidt van variatie versus retentie via exploratie versus exploitatie naar chaos versus orde en daarmee van een evolutietheoretische naar een chaostheoretische invalshoek. Kauffman legt dat verband expliciet: ‘Patching systems so that they are poised on the “edge of chaos” may be valuable (…)’ (1995: 129), evenals Volberda: ‘Maar tijdens het proces van blijvende strategische vernieuwing moeten oscillerende ondernemingen voorkomen dat ze ‘doorschieten’ en extreem rigide dan wel chaotisch worden.’ (1998: 73). Dat er een relatie bestaat tussen evolutietheoretische en chaostheoretische inzichten, is evident. De relatie komt in dit onderzoek wel aan de orde, de betreffende chaostheoretische inzichten zijn in dit onderzoek echter buiten beschouwing gebleven.
Veel systemen kennen vormen van aggregatie, vaak in de vorm van hiërarchische gelaagdheid. Zo kunnen in de biologie de organisatieniveaus molecuul, cel, weefsel, orgaan, organisme, populatie, soort, sociaal systeem en ecosysteem worden onderscheiden (Campbell 1974a: 179). Ook (menselijke) organisaties kennen een hiërarchische gelaagdheid die loopt van individu, via organisatorische structuren zoals teams, afdelingen en werkmaatschappijen, tot de organisatie als geheel. Bij deze aggregaties, en met name bij aggregaties in de vorm van een hiërarchische gelaagdheid, wordt hier aantal kanttekeningen gemaakt.
Aggregaties die uitsluitend het gemak van de analyse en de bespreking dienen, nominale aggregaties, moeten worden onderscheiden van aggregaties met eigen kenmerken, aanvullend aan die van de onderdelen waaruit ze zijn samengesteld. [8] Dit onderscheid is van belang omdat aggregaties zonder eigen kenmerken geen rol kunnen vervullen in variatie-, selectie- en retentieprocessen, anders dan in termen van de rol die de samenstellende delen daarin vervullen. [9] Waar in het vervolg over aggregaties wordt gesproken, worden uitsluitend niet-nominale aggregaties bedoeld.
Aggregatie is van belang omdat deze de variatie-, selectie- en retentieprocessen compliceert. Zo vindt biologische evolutie plaats op verschillende hiërarchische niveaus: het variatieproces op moleculair niveau, de selectie op het niveau van het individuele organisme, en de retentie weer op moleculair niveau. [10] Plaatsvervangende selectiecriteria (zie p. 18) vervullen een belangrijke rol bij het overbruggen van verschil in hiërarchisch niveau. Campbell spreekt van ‘a nested hiërarchie of selective-retention processes’ (1974c: 419). Ook March geeft aan dat deze gelaagdheid ook bij de bestudering van evolutionaire processen in organisaties complicerend werkt: ‘Units of adaptations are nested, so that some adapting units (e.g. individuals) are integral parts of other adapting units (e.g. organizations). (…) These features of organization considerably complicate any multilevel evolutionary story.’ (1994: 43).
Het in de beschouwing betrekken van hiërarchische lagen binnen een systeem noodzaakt tot formulering van de invloed die de lagen op elkaar kunnen hebben. Volgens de ‘starre’ of ‘rechtlijnige’ (Dennett 1995: 76) reductionistische visie zijn er twee reductionistische basisprincipes die betrekking hebben op deze gelaagdheid; ze worden door Campbell (1974a: 180) als volgt omschreven:
- alle processen die plaatsvinden op een hoger hiërarchisch niveau worden beperkt door en verlopen in overeenstemming met de wetten van de lagere niveaus, en
- de telenomische [11] prestaties op de hogere niveaus worden gerealiseerd door gebruik te maken van specifieke mechanismen en processen van lagere niveaus.
Aanvullend aan deze twee basisprincipes formuleert Campbell (ib.) twee aanvullende principes, die hij het emergentist principle en downward causation noemt. Het emergentist principle blijft verder buiten beschouwing, downward causation wordt door Campbell samengevat als een parafrase van het eerste basisprincipe: alle processen op een lager hiërarchisch niveau worden beperkt en verlopen in overeenstemming met de wetten van de hogere niveaus.
Over dit, in de ogen van starre reductionisten discutabele standpunt zegt Campbell: ‘Yet my position is not vitalist or teleological (…) since I advocate not the autonomy of the higher levels, but rather the additional restraints, aspects of selective systems that these higher levels encounter.’ (ib.: 182). Ook Dawkins, die door sommigen wordt gezien als een ‘starre reductionist’ onderschrijft dit, in een reactie op het de suggestie (van Lewontin) dat hij een genetic determinist is: ‘(…) it is perfectly possible to hold that genes exert a statistical influence on human behaviour while at the same time believing that this influence can be modified, overridden or reversed by other influences.’ (1989: 331).
Campbell’s downward causation en het verband tussen environmental change en enactment dat Weick beschrijft, hangen nauw samen. Waar Weick kiest voor een causaal model, zie Figuur 3-1, waarin de dynamiek van de verschillende onderdelen tot uiting komt, kiest Campbell voor een statisch model en komt de dynamiek pas in zijn beschrijving van downward causation naar voren: ‘The ‘causation’ is downward only if substantial extents of time, covering several reproductive generations, are lumped as one instant for purposes of analysis.’ (1974a: 180). Het ‘which then impose on them’ uit ‘the more active role (…) organizational members play in creating the environments which then impose on them.’ van Weick (1979: 130) komt zo overeen met het ‘restrained by’ uit ‘processes at the lower levels of a hierarchy are restrained by and act in conformity to the laws of the higher levels.’ van Campbell (1974a: 180).
In een van zijn laatste publicaties, How individual and face-to-face group selection undermine firm selection in organizational evolution, geeft Campbell (1994) een voor dit onderzoek relevant overzicht van inzichten op dit gebied. Een samenvatting van dit artikel is om die reden opgenomen als bijlage 3. Hier wordt volstaan met het opsommen van de in dit artikel genoemde bedreigingen voor het effectief verlopen van selectieprocessen. Op basis hiervan wordt een aanvullende voorwaarde aan het selectieproces geformuleerd, namelijk die van consistentie in de hiërarchie.
Selectiemechanismen op individueel niveau en op het niveau van primaire [12] groepen ondermijnen selectieprocessen op ondernemingsniveau. Dit is het gevolg van de potentiële conflicten die bestaan tussen 1) de belangen van de onderneming en die van ieder individu; 2) de belangen van een persoon en die van zijn of haar collega’s; 3) organisatorische niveaus in de onderneming; en 4) primaire groepen in de onderneming (ib.: 33). Van de problematiek rondom deze potentiële conflicten geeft Campbell een aantal voorbeelden:
- De van onderneming naar onderneming springende CEO en de betekenis die aan veranderingen wordt toegekend: ‘Too frequently the individual executive’s reputation is based upon his or her success in introducing a change (…) Note that success in introducing an innovation (…) is so generally regarded as a symptom of executive efficacy that a new head of a department or firm feels called upon to make changes whether needed or not, often destroying valuable practices and replacing them with inferior ones.’ (ib.: 34-35).
- Managementinformatiesystemen en de daardoor geïntroduceerde prestatiecriteria krijgen kritiek, omdat deze prestatiecriteria over het algemeen slechts proxy variables zijn, afgeleiden van de relevante prestatie: ‘Once any of such proxy variables becomes used in managerial control, “irrelevant” components that will produce the desired score come to dominate, and validity is undermined.’ (ib.: 36). In het verlengde hiervan ligt het spanningsveld dat in beursgenoteerde ondernemingen bestaat tussen korte- (CEO ) en langetermijn (ondernemings)-doelstellingen waarin de eerder genoemde proxy variables de effectiviteit van de onderneming ondermijnen: ‘It becomes essential for the CEO’s short-term job maintenance and bonus magnitude (individual selection) that these imperfect proxy variables of firm longevity, growth potential, and long-term profitability (firm-level selection) be kept at high readings, even at the expence of underlying long-term firm-level efficacy.’ (ib.: 37).
- De menselijke natuur en de naïviteit waarmee de werkelijkheid vaak tegemoet wordt getreden. Het concrete voorbeeld waarnaar hier worden verwezen betreft het invullen van formulieren waarmee gegevens wordt verzameld waaruit de hiervoor genoemde managementinformatie wordt afgeleid: ‘Once again [refererend aan een eerder citaat: ‘As naive experimental psychologists rather than cynical streetwise sociologists’], I suspect that the forms have been designed by world-unwise experimental psychologists who assume that the only motive in filling them out is to report accurately. Instead, our evolutionary analysis leads us to expect (…) that motives 2b [‘For the good of one’s own face-to-face group within the firm or the individuals within it’] and especially 2c [‘For one’s own well-being and that of one’s family’] will dominate.’ (ib.: 36).
Campbell besluit het artikel met de waarschuwing dat het gebruik van evolutietheoretische inzichten in de bedrijfskunde zich niet mag beperken tot de analogie tussen culturele evolutie en de selectieve overleving van ondernemingen, maar ook aandacht moet schenken aan de door de biologische evolutie gevormde menselijke natuur die leidt tot wat hij genetic competition among the cooperators noemt (ib.: 38).
De voorwaarde van in de hiërarchie consistente selectie houdt in dat de plaatsvervangende selectiecriteria congruent zijn met de externe selectiecriteria die zij vertegenwoordigen.
Figuur 3-4 geeft weer dat het variatie-, het selectie- en het retentieproces op zich kunnen worden beschouwd als evoluerende systemen, waardoor een hiërarchie ontstaat. De werkelijkheid is nog complexer: elk van de deelprocessen kan bestaan uit verschillende systemen die elkaar wederzijds beïnvloeden, en soms is het in analytische zin aanwezige onderscheid tussen, bijvoorbeeld, selectie en retentie afwezig, omdat deze deelprocessen overlappen of samenvallen.
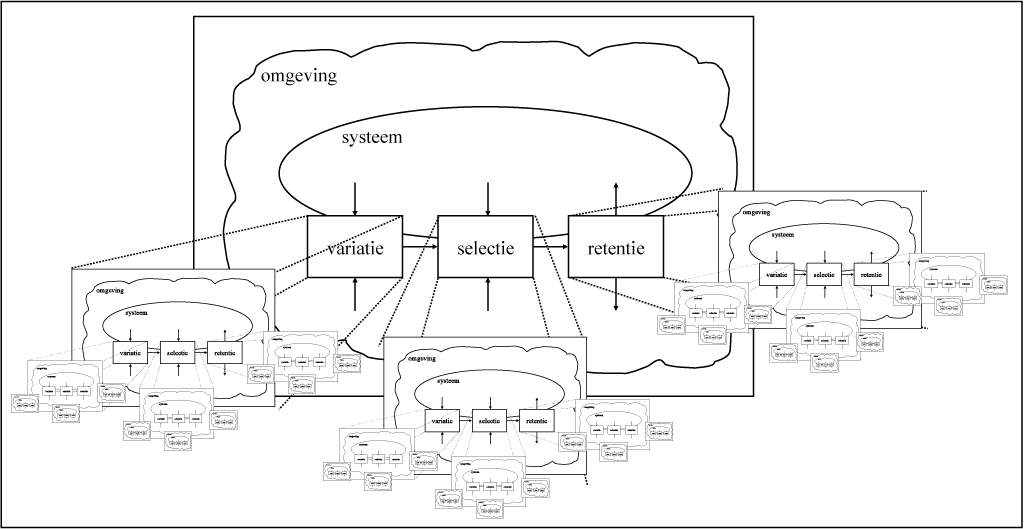
Figuur 3-4 Het Droste-effect van variatie, selectie en retentie
3.3 Evolutie: een algemeen model
Het algemene model van evolutie dat in deze rapportage wordt gehanteerd, is gebaseerd op de systeembenadering (Organisatiekunde 1A 1985, cursusdeel 2). Het formuleren van het algemene model in systeembenaderingstermen betekent dat (zie ook Figuur 3-5):
- het systeem en zijn omgeving zijn samengesteld uit entiteiten.
- het systeem wordt begrensd door de systeemgrens; de entiteiten binnen de systeemgrens behoren tot het systeem, de entiteiten buiten de systeemgrens behoren tot de omgeving van het systeem.
- entiteiten relaties met elkaar hebben; entiteiten binnen de systeemgrens kunnen relaties met elkaar en met entiteiten in de omgeving hebben, m.m. geldt hetzelfde voor de entiteiten in de omgeving.
- entiteiten en relaties eigenschappen hebben.
Het algemene model voor evolutie bestaat uit:
- Het evoluerende systeem: een open systeem, bestaande uit de entiteiten die tot het systeem behoren, de relaties tussen deze entiteiten, en de kenmerken van deze entiteiten en die van de relaties daartussen.
- De omgeving van het systeem, bestaande uit de entiteiten in deze omgeving, de relaties tussen deze entiteiten, en de kenmerken van deze entiteiten en de relaties daartussen.
- De relaties tussen het systeem en zijn omgeving, bestaande uit de relaties die betrekking hebben op groepen entiteiten waarvan er ten minste een behoort tot het systeem en ten minste een tot de omgeving van het systeem, en de kenmerken van deze relaties.
- Een evolutionair proces, in functionele zin bestaande uit de, in deze volgorde te doorlopen, deelprocessen variatie, selectie en retentie.
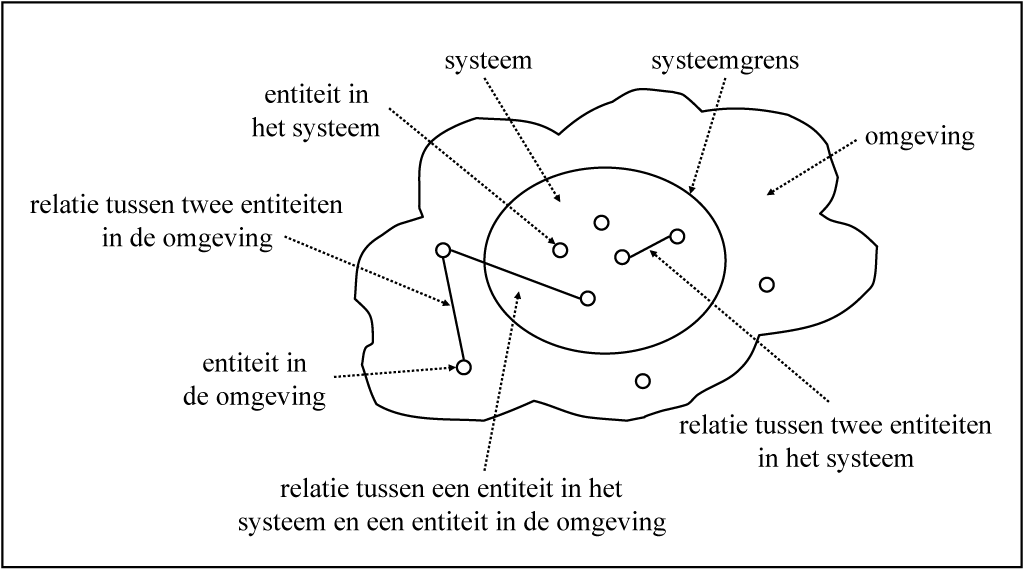
Figuur 3-5 Begrippen uit de open-systeembenadering
In de open-systeembenadering:
- kan de omgeving van het systeem dat object van onderzoek is, worden beschouwd als een verzameling systemen die in wisselwerking staan met het onderzochte systeem. Omdat deze systemen in de omgeving ook onderhevig zijn aan evolutionaire processen, is co-evolutie een verschijnsel dat impliciet onderdeel is van het algemene model.
- kunnen onderdelen van het systeem dat object van onderzoek is, worden beschouwd als systemen; ook deze subsystemen, waaronder die subsystemen die zorgdragen voor variatie, selectie en retentie, zijn onderhevig aan evolutionaire processen. Aggregatie en hiërarchische gelaagdheid maken daarmee impliciet onderdeel uit van het algemene model.
Het verschil tussen evolutie en natuurlijke selectie dat bestaat in de gebruikelijke biologische interpretatie van deze begrippen speelt in het algemene model geen rol, omdat een gegeneraliseerd mutatiebegrip als onderdeel van dit algemene model gedefinieerd kan worden, evenals een gegeneraliseerde vorm van genetische drift.
In het deelproces variatie ontstaan de variaties op basis waarvan selectie kan plaatsvinden. Het deelproces selectie geeft die variaties door aan het deelproces retentie die meer bijdragen aan een verbetering in de mate van afstemming tussen het systeem en zijn omgeving ten koste van variaties die dat in mindere mate doen. De vaststelling van de ordening van de bijdragen van de verschillende variaties vindt plaats op basis van selectiecriteria. Het deelproces retentie verzorgt de vastlegging in het systeem en in de omgeving van die variaties die het via het deelproces selectie krijgt aangeboden.
De variatie, de selectiecriteria en de vastlegging kunnen, gegeven de beschrijving van het systeem en zijn omgeving, betrekking hebben op:
- het systeem zelf:
- het aantal entiteiten waaruit het systeem is samengesteld,
- de kenmerken van de entiteiten,
- de relaties tussen de entiteiten,
- de kenmerken van de relaties tussen de entiteiten;
- de omgeving van het systeem:
- het aantal entiteiten in de omgeving waarmee een relatie bestaat,
- de kenmerken van deze entiteiten,
- de relaties tussen de entiteiten,
- de kenmerken van de relaties tussen de entiteiten;
- de relaties tussen het systeem en zijn omgeving:
- de relaties tussen groepen entiteiten waarvan er ten minste een behoort tot het systeem en een tot de omgeving en de kenmerken van deze relaties.
Door het evolutionaire proces ondergaat het systeem in de loop van de tijd wijzigingen. Beschouwen we dit proces als systeem, dan zijn de entiteiten T0, T1 enz. van dit systeem de toestanden waarin het evoluerende systeem zich op tijdstippen t0, t1 enz. bevindt, zie Figuur 3-6.

Figuur 3-6 Het evolutionaire proces: opeenvolgende toestanden van een systeem
Motivatie voor de gemaakte keuzen in model en schema
De keuze voor een modellering binnen de algemene regels van de open-systeembenadering is ingegeven door het frequente gebruik van aan deze benadering ontleende begrippen door zowel Campbell als Weick. De schematische weergave van omgeving en systeem zoals in Figuur 3-5 is in overeenstemming met het algemene gebruik om bij open systemen de omgeving expliciet weer te geven. De weergave van omgeving en systeem is in feite een Venn-diagram: de systeemgrens geeft aan welke entiteiten wel en welke niet tot het systeem behoren. Het feit dat entiteiten in het schema kunnen worden weergegeven, biedt tevens (beperkte) mogelijkheden om kenmerken van entiteiten, relaties tussen entiteiten en kenmerken van relaties tussen entiteiten schematisch weer te geven. De horizontale en verticale dimensie van het schema zijn betekenisloos.
Weick presenteert zijn model, zie Figuur 3-1, als een causaal model (Weick 1979: 69-88) . Figuur 3-1 geeft de relatie weer tussen vier processen die zich in organisaties afspelen en de pijlen die de verschillende processen met elkaar verbinden geven aan of er sprake is van versterking (+) of demping (−). De combinatie van de processen variatie, selectie en retentie is overgenomen uit het schema van Weick en toegevoegd aan het schema van Figuur 3-5 en deze vormen tezamen de schematische weergave van het algemene model van evolutie van Figuur 3 3. Daarmee bevat het schema van Figuur 3-3 twee lagen: een laag die gevormd wordt door de weergave van de entiteiten, verdeeld over het systeem en de omgeving en een laag die de processen weergeeft die zich afspelen.
Het deelproces variatie heeft relaties met de omgeving en met het systeem. Beide relaties worden weergegeven door een pijl in de richting van het deelproces variatie: in zowel de omgeving als het systeem vinden gebeurtenissen plaats die leiden tot het ontstaan van variaties. Ook het deelproces selectie heeft relaties met de omgeving en met het systeem, en ook deze relaties worden weergegeven door een pijl in de richting van het deelproces selectie: zowel de omgeving als het systeem beïnvloeden met hun selectiecriteria het selectieproces. De causale relaties die in het schema van Weick bestaan tussen retentie enerzijds en variatie en selectie anderzijds, zijn in dit model vervangen door de combinatie van de beïnvloeding van het systeem en de omgeving door het retentieproces (de uitgaande pijlen uit het retentieproces) en de invloed van het systeem en omgeving op het variatie- en selectieproces (de ingaande pijlen in het variatie- en selectieproces). In het model is evenmin de relatie opgenomen zoals die Weick legt van enactment naar ecological change omdat deze voor sociaal-psychologische processen wel, maar voor veel andere processen niet beargumenteerd kan worden. In plaats van een directe relatie van variatie naar omgeving biedt het algemene model de eerdergenoemde indirecte relatie, via selectie en retentie, naar de omgeving. (Het is mogelijk dat deze interpretatie overeenkomt met die van Weick omdat in zijn model geen directe relatie tussen retentie en omgeving voorkomt; wellicht wordt in zijn model de relatie van enactment naar ecological change uitsluitend gevoed door de relatie van retentie naar enactment.)
3.4 Voorwaarden voor effectieve ontwikkeling
Gegeven het algemene model voor evolutie kan de effectiviteit van het evolutionaire proces worden gedefinieerd als de verbetering in de mate van afstemming tussen het systeem dat door het evolutionaire proces wordt aangepast en zijn omgeving. Van de entiteiten T0, T1, T2 enz. in Figuur 3-6 kan de mate van afstemming A0, A1, A2 enz. tussen systeem zijn omgeving worden vastgesteld, waarna voor de periode t0 - t1 de absolute en de relatieve verbetering kunnen worden bepaald: (A1 - A0) en (A1 - A0) / A0.
De beschikbaarheid van het algemene model van evolutie en de definitie van de effectiviteit van het evolutionaire proces laten onverlet dat de parameters die de waarde van de mate van afstemming tussen een systeem en zijn omgeving bepalen verschillen voor de vele verschillende systemen die bestudeerd kunnen worden. Met andere woorden: er is wel een algemeen model, maar er is geen algemene interpretatie van de werkelijkheid mogelijk op basis van dat model. De bioloog John H. Campbell is het met deze zienswijze voor zijn eigen vakgebied niet onverkort eens: in tegenstelling tot veel van zijn collega-biologen stelt hij evolutie gelijk aan vooruitgang. Hij verwoordt in het volgende citaat zowel de gangbare mening binnen zijn domein als zijn eigen mening:
It seems to me that several contributors to this volume overlook this view of the progression in evolution because they imagine that progress must be judged by a criterion outside of evolution, such as fitness or complexity. As they point out, the choice of any external yardstick for defining advancement is arbitrary. Elegant simplicity is just as valid as messy complexity for rating organisms. Obviously, the appropriate measure for evaluating evolutionary progress must be life’s ability to evolve. (1994: 100).
In de cursus Organisatieontwerp wordt het als volgt onder woorden :
Men kan de contingentietheoretische denkwijze samenvatten in een eenvoudig model. De effectiviteit van een organisatie wordt bepaald door de mate van afstemming of ‘fit’ tussen de structuur van de organisatie en de situatie of context waarin de organisatie zich bevindt. Men noemt dit de congruentie- of fit-hypothese. (…)
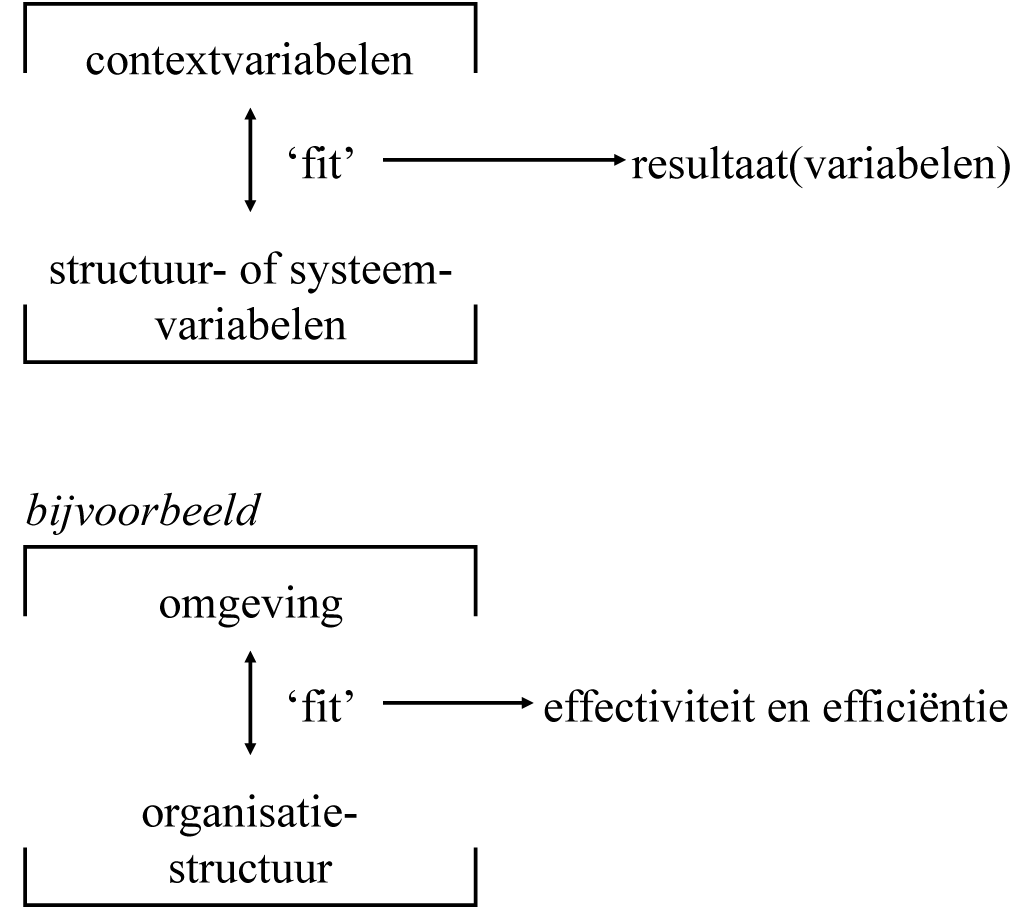
Wat men in die context echter (precies) van belang acht voor het functioneren van organisaties, en hoe men begrippen als omgeving, technologie, structuur en resultaat of effectiviteit definieert, verschilt nogal per auteur. (Organisatieontwerp 1991, cursusdeel 1: 107).
Alvorens dieper in te gaan op de voorwaarden voor effectieve ontwikkeling, is het van belang te constateren dat aan veel systemen impliciet een continuïteitsdoelstelling wordt toegedacht. Voor bepaalde categorieën systemen is dat terecht, zo hebben veel organisaties een doelstelling waarvan continuïteit impliciet of expliciet onderdeel uitmaakt. Voor veel andere systemen echter is het overleven een doelloos effect van de werking van de evolutionaire processen; de populatie veldmuizen in de provincie Utrecht is hier een voorbeeld van.
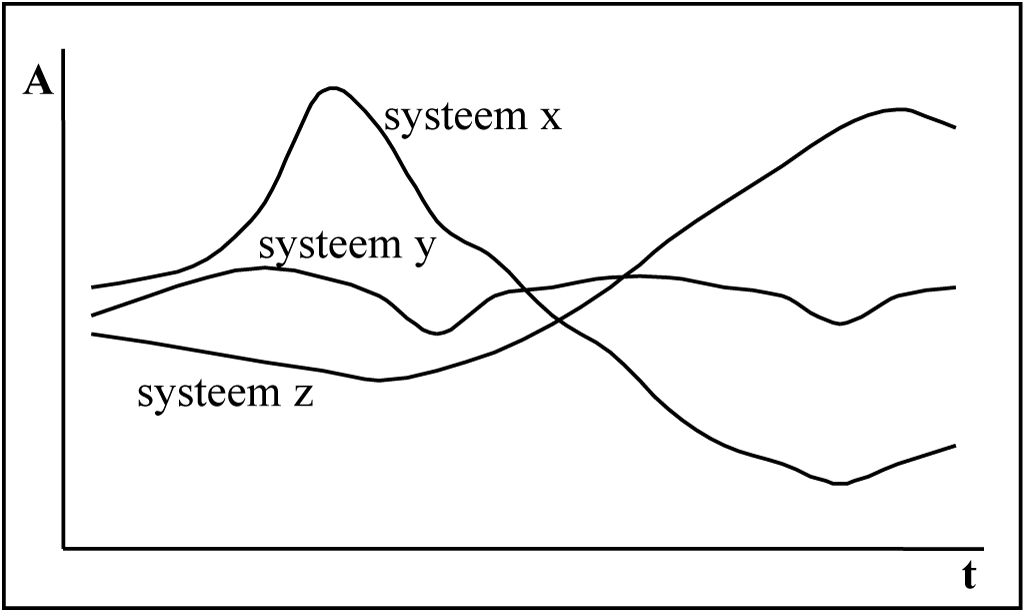
Figuur 3-7 Verbetering in de mate af afstemming?
Dit aspect continuïteit compliceert de operationalisatie van het begrip ‘effectieve ontwikkeling’ in ‘verbetering in de mate van afstemming tussen het systeem en zijn omgeving’. Dit wordt gevisualiseerd in Figuur 3 7, en door March als volgt onder woorden gebracht voor een bepaalde discipline:
Engineering traditions have ordinarily treated the definition and measurement of improvement as unproblematic. In fact, of course, improvement is difficult to define and measure, and every experienced engineer knows it. (…) The history of engineering is cluttered with examples of steps that seemed to offer short-run improvements in some domains but in retrospect were deemed to have created greater problems in the long run or in others domains. (March 1994: 46).
Uit het algemene model en uit de literatuur waarop dit model is gebaseerd kunnen voorwaarden voor effectieve ontwikkeling worden afgeleid. Primair is het actief aanwezig zijn van de deelprocessen, variatie, selectie en retentie. ‘Lacking any one of these, no increase in fit or order will occur. The occurence of all three is a rare event—fit and order are rare events.’ (Campbell 1974b: 143). Aan de drie deelprocessen worden aanvullende eisen gesteld, wil er sprake kunnen zijn van effectieve ontwikkeling; deze eisen zijn reeds genoemd bij de beschrijving van de processen in § 3.2 en worden hier geresumeerd.
Aanvullende eisen aan het deelproces variatie:
- ‘blind and multiple variations with failures’ (Romanelli 1999: 81):
- blind: het proces dient te resulteren in variaties die op geen enkele wijze zijn afgestemd op een verwacht resultaat.
- multiple: uit het variatieproces dienen meerdere variaties te resulteren.
- with failures: tot de door het variatieproces geproduceerde variaties dienen ook mislukkingen te behoren.
- ‘persistence until natural selection can operate’ (Weick 1979: 129): de door het variatieproces afgeleverde variaties dienen lang genoeg te blijven bestaan om door het selectieproces verwerkt te kunnen worden.
Aanvullende eisen aan het deelproces selectie:
- consistent selection criteria’ (Campbell 1965: 27):
- consistentie in de tijd: selectiecriteria dienen relatief onveranderlijk te zijn over een langere periode.
- consistentie in een hiërarchie van selectiecriteria: selectiecriteria op een lager niveau moeten de selectie op een hoger niveau niet ondermijnen.
Aanvullende eis aan het deelproces retentie:
- ‘liability to recall’ (James, geciteerd in Weick 1979: 207): de wijze waarop retentie plaatsvindt dient het mogelijk te maken het resultaat daarvan in het evolutionaire proces te gebruiken.
Ten slotte dient er, als vastgesteld moet kunnen worden of er sprake is van een effectieve ontwikkeling, een kwantificeerbare definitie van effectiviteit te zijn. Op basis van deze kwantificeerbare definitie kan operationalisatie plaatsvinden, met andere woorden: kan gemeten worden in hoeverre er, gegeven de definitie, sprake is van een effectieve ontwikkeling.
Terugkoppeling naar de probleemstelling
In dit hoofdstuk is deelvraag 1 beantwoord: uit de beschikbare literatuur is een algemeen model voor evolutie afgeleid. In § 3.3 is dit beschreven, waarbij de verschillende evolutionaire processen zijn benoemd, en verantwoording is afgelegd over de gemaakte keuzen bij de formulering ervan. In § 3.4 zijn de voorwaarden voor effectieve ontwikkeling opgesomd.
voetnoten bij hoofdstuk 3:
[1] In het vervolg wordt regelmatig aan de biologische context van het evolutiebegrip gerefereerd; zie eventueel bijlage 1 voor een toelichting.
[2] Campbell’s loopbaan was bijzonder omdat hij enerzijds als onderzoeker, bench scientist, veel publiceerde, maar daarnaast ook schreef over methodologie en kennisleer. Voor zijn eerste publicatie over kennisleer in 1959 had hij al vijftig onderzoekspublicaties op zijn naam staan. Ook toen de nadruk in het aantal publicaties kwam te liggen op het onderwerp evolutionaire kennisleer, bleef hij publiceren als onderzoeker. Deze ongebruikelijke combinatie van onderzoek en filosofie leidt tot een zekere verwevenheid; zo is de eerste publicatie van zijn generieke dogma in 1960 te vinden in het specialistische tijdschrift Psychological Review (Baum & McKelvey 1999: 1-2).
[3] Zie bijlage 2 voor een toelichting op het feit dat ‘evolutie’ een beladen begrip was, en lange tijd taboe in de sociale wetenschappen.
[4] De visie van Campbell werd door Popper geaccepteerd: ‘For me, the most striking thing about Campbell’s essay is the almost complete agreement, down even to minute details, between Campbell’s views and my own.’ (Popper 1974: 1059).
[6] Dat blijkt behalve uit de vele referenties in Weick (1979) aan Campbell ook uit de acknowledgements in het hiervoor genoemde boek: ‘Three others who supported and influenced the volume you are holding, but, for one good reason or another, are not represented directly are (…) and Karl Weick (Baum & McKelvey 1999: xii).
[7] De aandacht voor kunstmatige, in tegenstelling tot natuurlijke, selectie, is ook aanwezig bij Darwin en Campbell. Voor Darwin zijn de resultaten van kunstmatige selectie een bron van inzicht in het ontstaan van variatie (1859: 71-113) . Voor Campbell is kunstmatige selectie een via natuurlijke selectie verworven vaardigheid, een shortcut process, zie zijn op p. 13 beschreven dogma. De door managers van organisaties in het leven geroepen planningsprocedures, controlemechanismen enz. zijn zulke shortcuts, en maken van organisaties ‘the environment of the breeder’. Kennis van de werking van het proces van natuurlijke selectie leidt dan tot een bewust gebruik van een kunstmatige vorm daarvan: bewust variatie aanbrengen, beoordelen wat wel en niet werkt, en proberen het goede te behouden: cultiveren.
[8] Campbell (1990: 4) spreekt hier van ‘ontologically “real” levels of organization’ en schaart zich (1994: 24) aan de zijde van de sociologen die stellen ‘Groups are real!’
[9] Zo kan in de biologische evolutie geen selectie plaatsvinden op soortniveau omdat een soort geen kenmerken heeft, anders dan die van het totaal van de organismen waaruit deze bestaat.
[10] Dit is een simplificatie: in werkelijkheid vinden ook op moleculeniveau variatie-selectieretentieprocessen plaats. Dit is echter niet relevant in de context van dit onderzoek.
[11] Campbell (1974a: 180) gebruikt het begrip teleonomy, en later (1990: 4) het begrip telenomy: ‘For most of us teleological causation is rejected, but purpose-like achievements in biological organisms are recognized. In accordance with many biologists, the term telenomy (…) can be used to designate our position.’
[12] Face-to-face groups werden in de Angelsaksische literatuur voorheen primary groups genoemd. In het Nederlands is ‘primaire groep’ de gangbare vertaling van primary group, en ontbreekt een vertaling van face-to-face group. Daarom wordt hier ‘primaire groep’ gebruikt als vertaling van zowel face-to-face group als primary group. Inhoudelijk is dit geen bezwaar: onder al deze begrippen wordt in het kader van dit onderzoek verstaan: een groep die regelmatig persoonlijk contact heeft.
Wil Leeuwis, doctoraalscriptie bedrijfskunde, juni 2002
